长期以来,植物代谢酶被认为仅定位于细胞质、线粒体或质体中,作为“专职催化剂” 负责维持细胞的能量代谢与物质合成。然而,近年来的研究揭示,这些传统的代谢酶能够响应发育信号和环境胁迫,从其经典的亚细胞定位穿梭至细胞核,并在此获得全新的调控功能,即 “Moonlighting”(兼职)现象。目前,植物中核定位代谢酶的调控机制、作用模式及其生物学意义仍缺乏系统性的概念框架,这已成为植物代谢与表观遗传调控交叉领域的关键前沿科学问题。
近日,广西大学农学院徐秋涛教授、张积森教授团队在Cell Press旗下生物化学与分子生物学领域权威期刊Trends in Biochemical Sciences发表了题为“Stimulus-Responsive Nuclear Moonlighting of Plant Metabolic Enzymes Rewires Chromatin”的综述文章。该研究系统总结了植物代谢酶核转位现象的最新进展,创新性地提出了代谢酶通过三种非互斥模式连接代谢与染色质调控的概念框架,并指明了该领域未来亟待解决的关键科学问题,为深入理解植物代谢—表观遗传修饰的相互调控关系提供了重要理论框架与全新视角。

在本文中,作者首先梳理并整合了多个经典的代谢酶核定位事件。结果显示,植物激素乙烯刺激、光照变化、高温胁迫以及氧化胁迫等信号,能够分别诱导丙酮酸脱氢酶复合体(PDC)、丙酮酸激酶(PK)、α-酮戊二酸脱氢酶(KGDH)以及甘油醛-3-磷酸脱氢酶(GAPDH)等多种核心代谢酶发生精准的核质转运。进一步对这些核内代谢酶的作用机制进行归纳分析,作者提炼出三种功能模型:代谢物供应模型,即核内酶为染色质修饰提供关键底物。例如ACLA2与PDC在核内产生乙酰辅酶A等;酶—染色质修饰因子互作模型,即代谢酶直接与组蛋白修饰酶形成功能性复合物。例如KGDH与JmjC去甲基化酶互作、MBP-1与SRT1协同调控;以及直接染色质修饰调控模型,即代谢酶自身演化为非经典的组蛋白修饰酶,直接催化组蛋白化学修饰,例如PK1和PK6-8直接磷酸化组蛋白H3T11。这些发现共同揭示:核定位代谢酶已从被动的底物供应者,跃升为主动的“兼职染色质调控因子”。
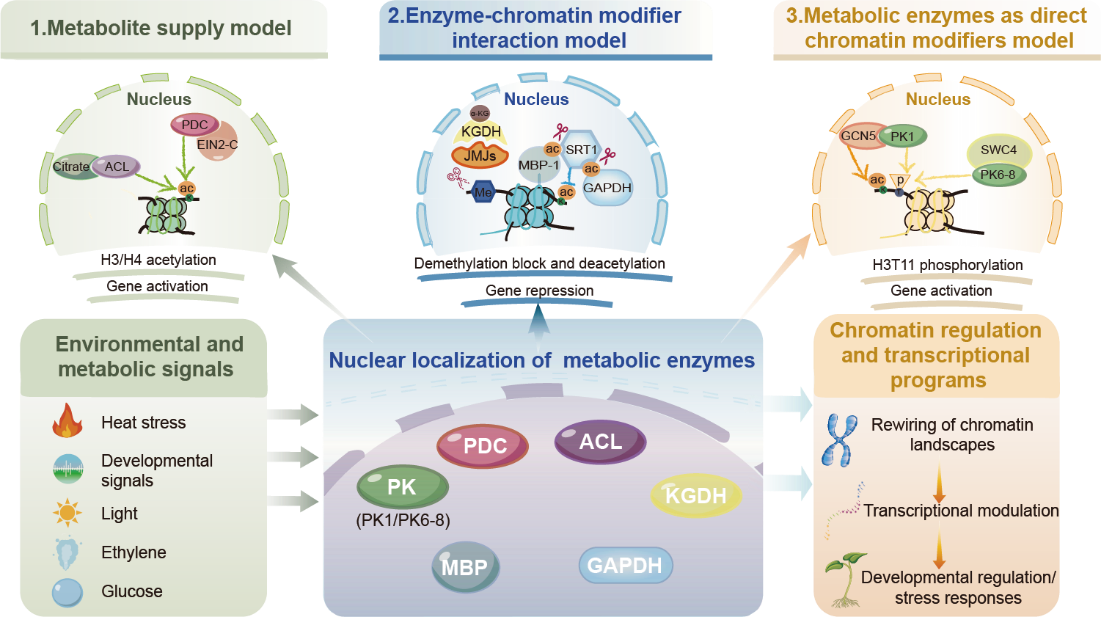
图1代谢酶通过三种非互斥模式连接代谢与染色质调控的概念框架
此外,文章还将这一视角拓展至跨物种层面,指出从哺乳动物到植物,利用代谢酶作为核内表观遗传调控因子的策略在进化上高度保守,形成了一条贯穿“代谢—染色质”的信息桥梁。不过,作者也明确指出,目前该领域仍面临多重挑战:仅有部分酶的刺激响应性核转位获得了严格的内源验证;核内直接功能与全局代谢重编程的间接效应尚难以精准区分;植物中潜在的核内代谢酶全谱、引导其精确定位的信号分子、以及实现靶基因位点特异性调控的分子机制,仍是亟待回答的关键科学问题。
综上所述,该文系统构建了植物代谢酶通过核内“兼职”功能连接代谢与表观遗传调控的概念框架,不仅深化了对植物发育可塑性及环境适应机制的理解,更为直接操控代谢—表观遗传通路以精准设计抗逆、高产作物的未来育种策略,提供了新的概念基础和潜在靶点。
广西大学为该论文第一完成单位,广西大学农学院硕士生肖贵毓为第一作者,广西大学农学院徐秋涛教授与张积森教授为共同通讯作者。该研究获得广西杰出青年科学基金、广西科技重大专项、国家自然科学基金等项目资助。
编辑 | 韦星璇
一审一校 | 黄婧
二审二校 | 王邕
三审三校 | 张积森